小的时候有没有玩过一个游戏,一张纸可以对折几次?在目前的世界纪录中,一张纸最多被折叠了13次,高达1.6米的高度。在人体的细胞里有近2米长的DNA,却要被紧凑地塞进直径不到10微米的细胞核中,你会怎么折?别担心,细胞早已雇用了三位专业“打包工”:
打包工 |
主要功能 |
环环——Extrusive Cohesin(挤压型黏连蛋白) |
如“织布工”般推出DNA环,构建染色质拓扑结构域(TADs),为DNA复制铺设框架 |
粘粘——Cohesive Cohesin(连接型黏连蛋白) |
在DNA复制后充当“胶水”,将姐妹染色单体粘连,确保染色体正确分离 |
大力——Condensin(凝缩蛋白) |
将染色质螺旋化压缩成棒状染色体,让DNA高效进入分裂阶段 |
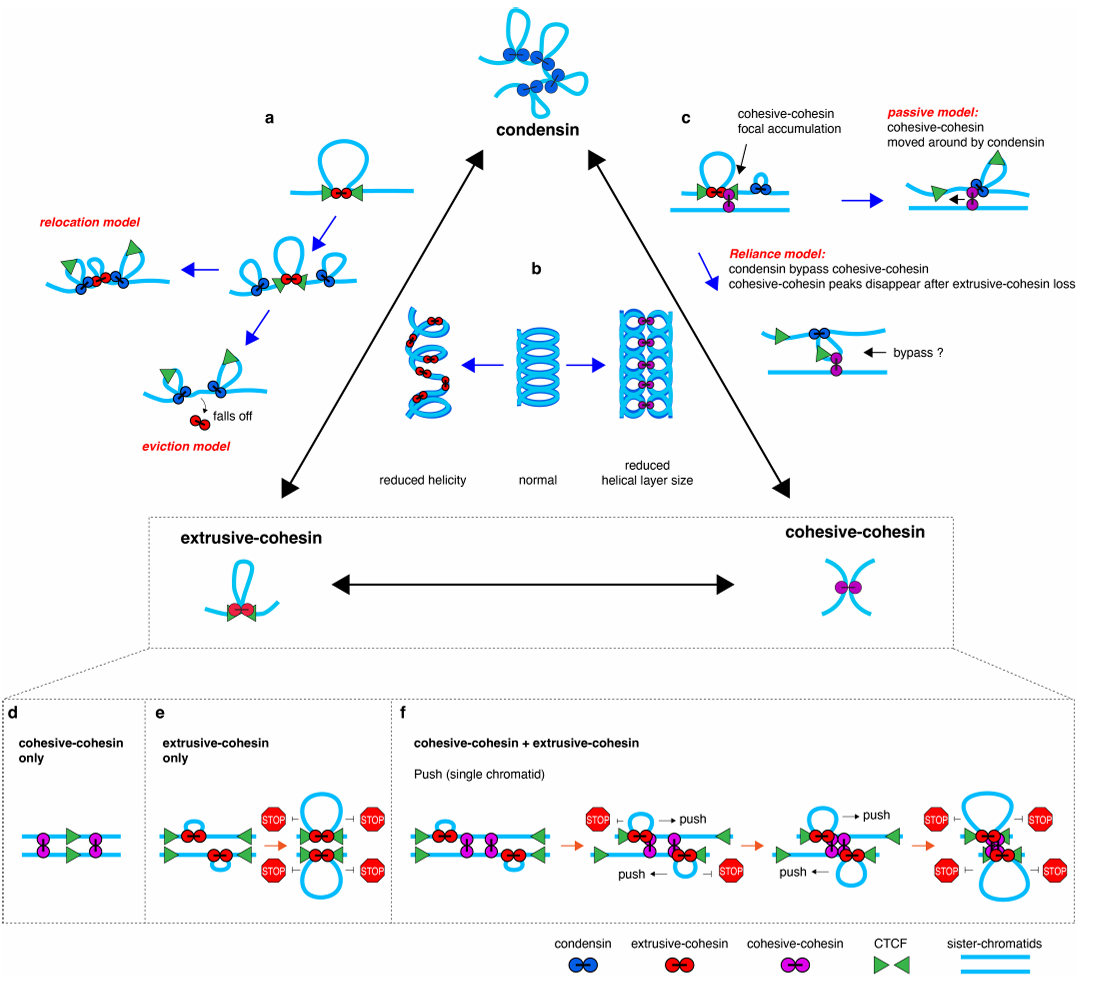
尽管三者功能已知,它们如何在时空上交叠协作却困扰学界十余年。例如,在细胞分裂前的黄灯期,三者同时存在——环环和粘粘尚未退场,大力已准备登场。此刻它们如何避免“交通堵塞”?两种黏连蛋白的是否缺一不可?这些问题直接关联基因组稳定性,异常则导致癌症或发育疾病。经过SZBL科学家们研究发现:这些打包工可不是轮班制,而是同场竞技,博弈不断!
2025年2月,SZBL张浩岳团队携手黄恺团队,在Nature 在线发表了最新研究,题为:Extensive mutual influences of SMC complexes shape 3D genome folding(张浩岳团队揭示三种SMC复合物如何协同调控染色质折叠)。他们开发出一套“三重可控降解系统”,首次在G2/M期阶段实现了对三种SMC蛋白——环环(extrusive cohesin)、粘粘(cohesive cohesin)、大力(condensin)的独立操控,并通过Hi-C、ChIP-seq、显微成像和理论模拟,全景揭示了染色质折叠的动态机制。
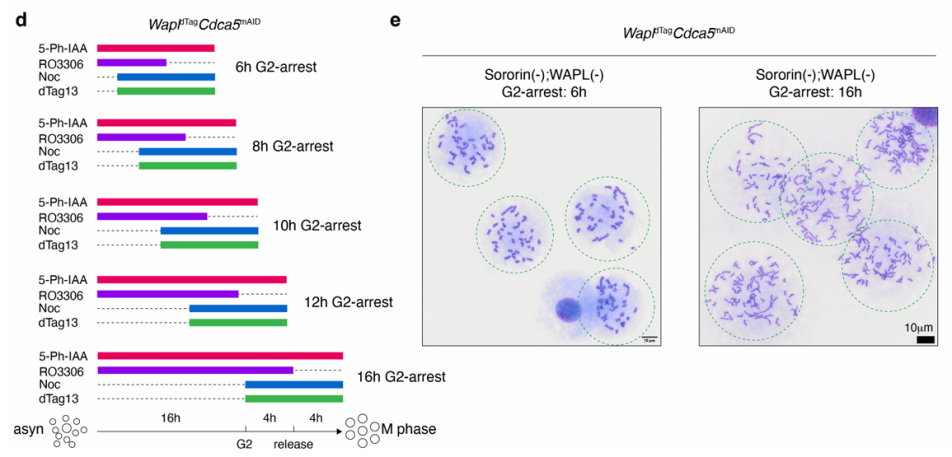
该项研究所采用的三重诱导降解系统
本研究通过正交降解系统(dTag靶向WAPL,mAID靶向Sororin或SMC2)与G2/M阻滞-释放策略(RO-3306阻滞后药物时序处理)的协同设计,首次在有丝分裂染色体上实现了三种SMC复合物的独立操控。这一核心方法解决了它们在S期后天然共存的难题,为解析其动态互作提供了精准的实验体系。
研究结果:三大“打包工”同台上岗,谁协作?谁掣肘?
Step 1:大力“清场”,环环的精致结构全线崩盘
在细胞尚未分裂时,环环(extrusive cohesin)像个折纸达人,精细地把DNA这根长线折叠成一个个有规则的小圈——这些染色质环(loops)一个接一个地排列起来,最终形成了结构分明的街区,叫做TADs(拓扑关联域)。TADs就像城市里的功能分区,把基因活动划分得井井有条。折叠的“停靠点”往往由CTCF这位“交通指挥员”设定,他能指引环环在特定位置停下,精准塑造基因组架构。但一到M期,大力(condensin)闪亮登场。他的任务不是维持秩序,而是大扫除!要让染色体快速压缩、整装待发,就得先拆掉这些圈和街区,清空旧结构,为新的折叠方式腾出空间。
张浩岳团队发现,大力一加入,环环辛苦打造的TADs立刻土崩瓦解。Hi-C图谱显示,TAD边界变得模糊甚至完全消失;ChIP-seq结果也证实,RAD21(环环的核心成分)在CTCF等锚点上的信号显著下降。更有意思的是,大力对另一位打包工粘粘(cohesive cohesin)也有类似的清理效果,说明他有一套统一的“清扫流程”,为染色质压缩打好前战。
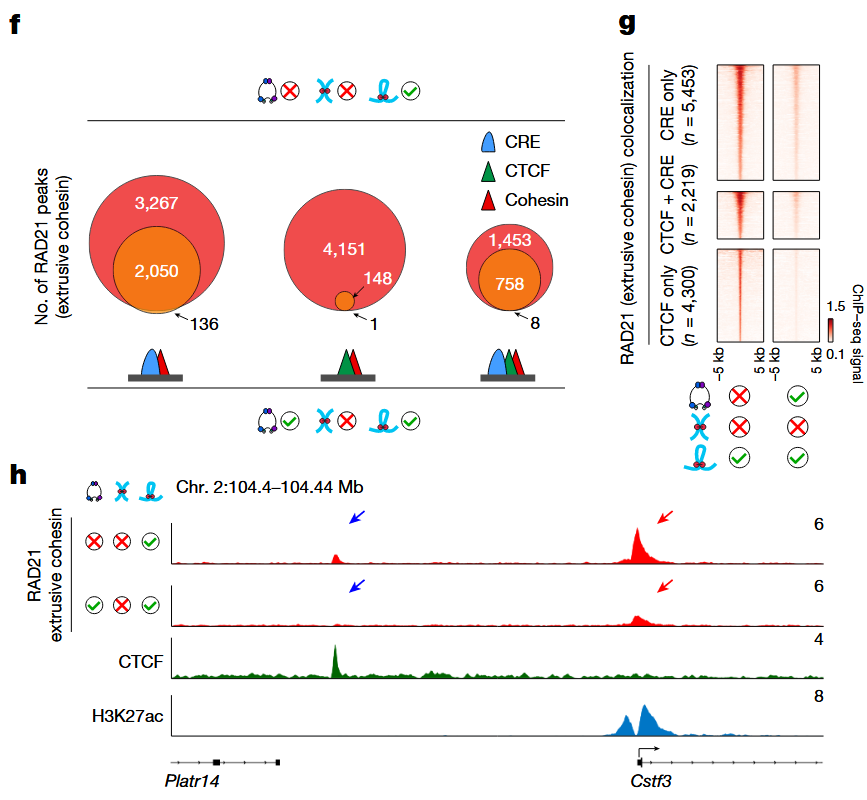
大力清除环环的基因组定位
Step 2::环环虽败犹荣,减速挡压做指挥
别看环环在大力面前败下阵来,他可没退出战场,而是悄悄换了打法。当大力独自作战时,染色质像弹簧一样迅速压缩。但只要环环还在,他就像给这跟弹簧套上了一圈圈缓冲垫,显著减缓了压缩节奏。更厉害的是,环环还成了幕后“指挥官”——他会主动把粘粘带到特定位置,一起抵抗大力的压缩推进。
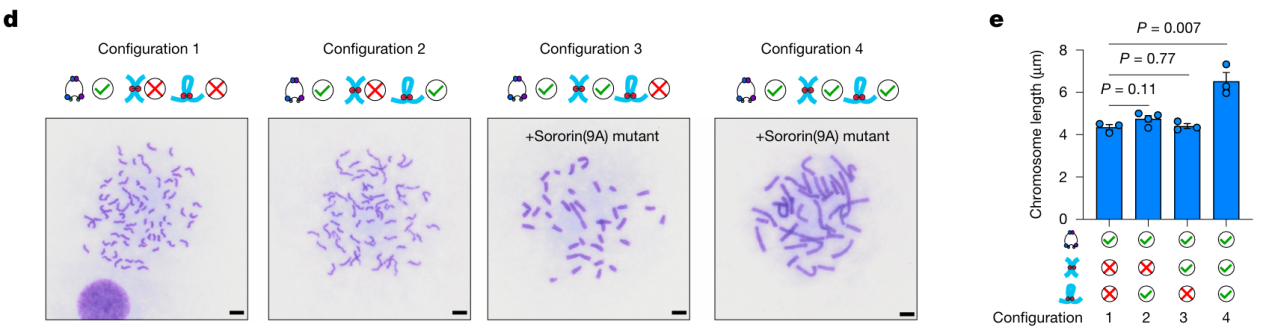
三位打包工不同配置下的染色体形态比较
配置1:仅大力(极度紧缩);配置2:大力+环环(明显舒展);配置3:大力+粘粘(轻微舒展);配置4:大力+环环+粘粘(最舒展状态)
Step 3:粘粘低调上线,幕后掌握折叠尺寸
粘粘虽然没有挤环能力,也不会挂靠在CTCF上,更构不出TADs结构,但可千万别小看他的作用。单打独斗时,粘粘几乎无法主导染色体结构的变化。但只要环环一出现,他立刻变身为“结构调节器”——控制环的扩展范围,避免DNA折得过紧、卷得过长。如图4显示,粘粘能优先抑制那些锚点弱、环大、RAD21信号低的结构,对锚点强、RAD21集中的区域几乎不干扰。这说明他能进行选择性抑制,是折叠节奏里的隐形稳定侠。
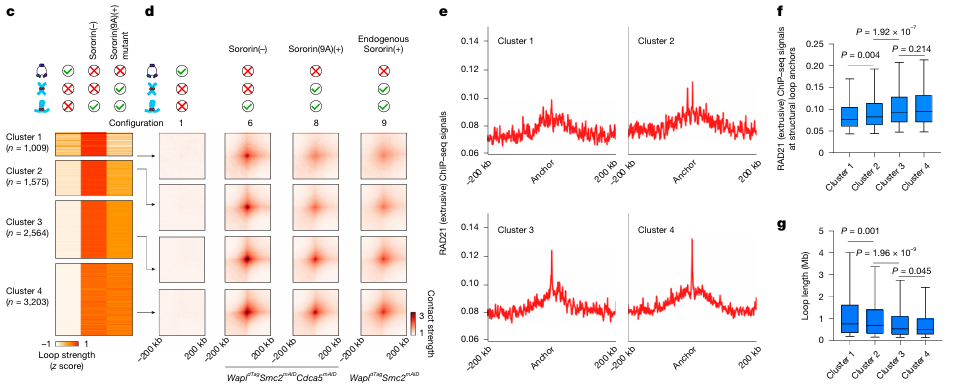
粘粘对染色质环的动态调控作用
Step 4:环环遇上粘粘,推还是停?模拟来揭晓
虽然实验揭示了环环与粘粘联手可以限制大力的压缩行为,但当粘粘与环环彼此“相遇”时,会发生什么?是撞停?绕行?还是直接推过去?为了回答这个问题,张浩岳团队找到了同在SZBL的黄恺课题组进行合作,构建了三种可能的模型:
①停滞模型(stalled):环环被粘粘阻挡,无法前进;
②绕行模型(bypass):双方各走各路,互不干扰;
③推挤模型(push-through):环环直接将粘粘推着走。
研究团队对每种模型下形成的环结构进行模拟,并与实验获得的Hi-C数据进行对比后,发现“推挤模型”与真实结果最吻合!结果显示:环环在推进时会把粘粘一路推着向前走,直到CTCF才停下。而这一过程不会干扰到另一条染色单体。这个过程不仅解释了loop的形成机制,也说明了粘粘限制loop扩展的能力从何而来。
这项研究首次揭示了三种SMC复合物在细胞分裂前同台竞争的动态互作模式。他们不是各司其职、互不干涉,而是在分裂前夕展开了一场错综复杂的“空间调控博弈”:
大力清除原有结构,开启压缩;
环环一边布阵,一边缓压;
粘粘低调控场,掌控折叠尺度。
三种SMC复合物的相互作用模式图
三者的角力,构建了染色体从松散到致密的折叠转变过程。而这场“平衡游戏”一旦被打破,就可能引发严重的染色体异常。事实上越来越多的研究表明,染色体异常折叠与多种人类疾病密切相关,比如:
而本文对于SMC复合物“互相掣肘又彼此协作”的动态机制的揭示,正为我们提供了一把重新理解这些疾病发生机制的钥匙。是否正是由于SMC三者之间的互作失衡,导致染色体环结构紊乱、折叠方向错误,最终引发结构性变异(如染色体缺失、易位)?这也引出了张浩岳团队更进一步的研究方向:在活细胞中监测三位打包工的现场操作,为设计药物精准修复打包事故提供依据。
每一次精妙的折叠,都是细胞精密的时空计算;每一次默契的协作,都是生命系统高度组织的体现。科学的意义,从不是把世界变复杂,而是用理解,为生命争取更多可能。
在这个神秘的细胞“折叠工地”里,谁是你心中的“最佳打包工”?

欢迎在评论区投票告诉我们你pick谁!



 粤公网安备44031102000926号
粤公网安备44031102000926号